




用户文章 IF=13.6|AHR介导的 m6A RNA甲基化是 PM2.5 诱导斑马鱼幼体心脏畸形的原因之一
发布时间:2024-03-13 14:40 | 点击次数:

越来越多的证据表明,暴露于环境中的细颗粒物(PM2.5)会抑制心脏发育,但其潜在机制仍然难以捉摸。2023年9月,云序客户苏州大学陈涛团队在Journal of Hazardous Materials(IF=13.6)上发表题为“AHR-mediated m6A RNA methylation contributes to PM2.5-induced cardiac malformations in zebrafish larvae”的研究性论文。在这项研究中,通过m6A Merip-seq、 RNA-seq 等实验证明了 PM2.5 中的可萃取有机物(EOM)会显著降低斑马鱼幼体心脏中全局 m6A RNA 甲基化水平,而甲基供体甜菜碱则可恢复这一水平。甜菜碱还能减轻 EOM 诱导的 ROS 过度生成、线粒体损伤、细胞凋亡和心脏缺陷。此外,作者发现 EOM 激活的芳基碳氢化合物受体(AHR)直接抑制了甲基转移酶 mettl14 和 mettl3 的转录。EOM 还诱导了全基因组 m6A RNA 甲基化变化,还发现,EOM上调了traf4a和bbc3这两个凋亡相关基因的表达水平,但通过强制表达mettl14,它们的表达水平又恢复到了控制水平。
该研究中的m6A MeRIP-seq 、RNA-seq 高通量测序均由上海云序生物公司提供。
研究背景
环境细颗粒物(PM2.5)是影响全球健康的一个主要风险因素,很容易进入循环系统并穿过胎盘屏障,对胎儿发育造成不利影响。先天性心脏病(CHD)是人类最常见的先天性缺陷,约占所有活产婴儿的 1%。先天性心脏病的病因尚不清楚,但目前认为是由遗传和环境因素共同作用造成的。多项流行病学研究表明,孕期暴露于 PM2.5 与后代的先天性心脏病密切相关。动物研究也表明,暴露于 PM2.5 会显著增加小鼠、鸡和斑马鱼的心脏畸形率。然而,PM2.5 的心脏发育毒性的内在机制仍有待阐明。
研究结果
(1)甲基供体甜菜碱减轻了EOM诱发的心脏缺陷并减少了m6A甲基化
我们首先检测了EOM中16种EPA优先考虑的多环芳烃的浓度,并确定了其中9种的存在。BaP当量为25.9纳克/立方米。由于采样期间 PM2.5 的平均水平为 66.5 微克/立方米,因此本研究中 EOM 的暴露浓度(5 毫克/升)大致相当于 1.95 微克/升 BaP,这是一个与环境相关的浓度水平。然后,我们研究了甲基供体甜菜碱对 EOM 的心脏发育毒性的影响。如图 1A 和 B 所示,补充甜菜碱可显著减轻 EOM 引起的心脏畸形(即心包水肿、心脏变长)和心功能障碍(表现为心跳率下降和 QT/RR 间期延长)。此外,EOM还明显降低了斑马鱼幼体心脏中m6A RNA的全局甲基化水平,而加入甜菜碱后,这一水平又恢复到了控制水平(图1C)。
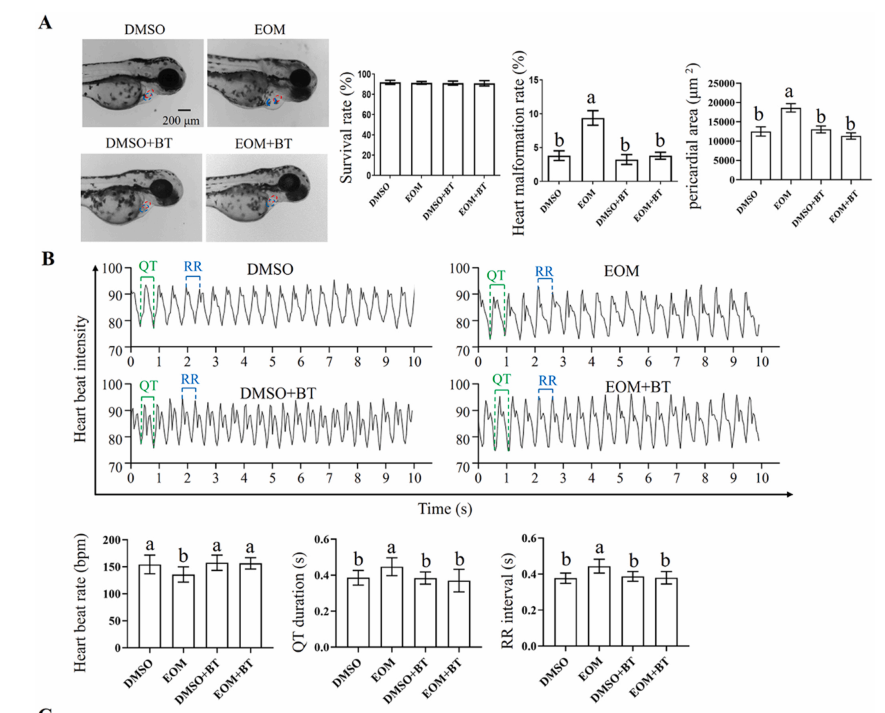
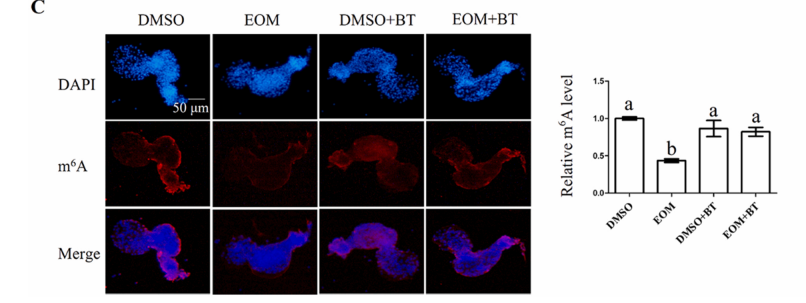
图 1. 72 hpf 时斑马鱼幼体的心脏缺陷和总体 m6A 水平(n ≥ 5)
(A)72 hpf 时斑马鱼幼体的代表性图像,以及胚胎存活率、心脏畸形率和心包面积的量化结果。虚线环绕心房(蓝色)或心室(红色)
(B)心跳强度和量化结果。bpm:每分钟心跳次数;QT:Q 波和 T 波之间的间隔;RR:R 波之间的间隔。
(C)m6A 甲基化水平的代表性图像和量化结果。EOM:浓度为 5 mg/L 的 EOM;BT:浓度为 5 μM 的甜菜碱。不同字母表示差异显著。
(2)甜菜碱可减少 EOM 诱导的 ROS 生成、MMP 崩溃和细胞凋亡
如图2A和图B所示,EOM显著增强了斑马鱼幼体心脏细胞内ROS和mtROS的产生,而添加甜菜碱则可抵消这种作用。甜菜碱还减轻了EOM诱导的氧化应激相关基因(包括nrf2a、sod2、cat、gstp1和gstp2)的过表达。此外,EOM降低了斑马鱼胚胎心脏中MMP的水平并增强了细胞凋亡,而补充甜菜碱后这两种情况都得到了恢复(图2 C和D)。
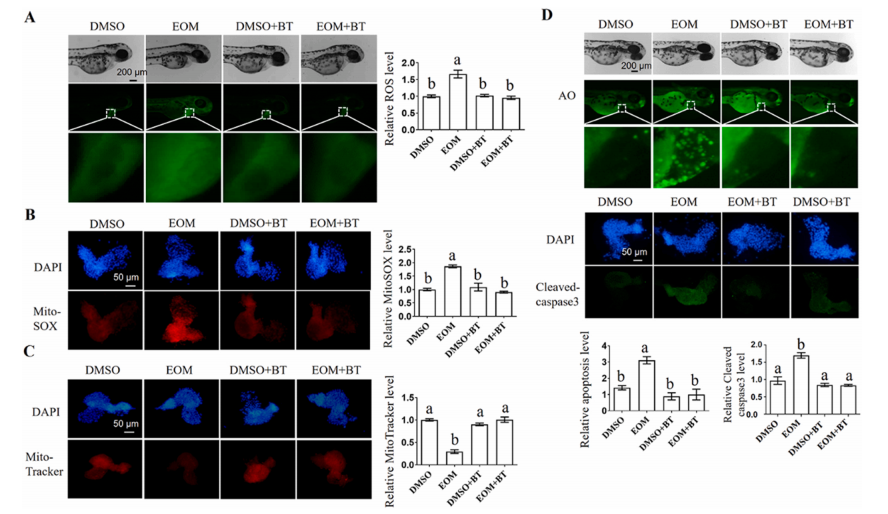
图2. 72 hpf 时斑马鱼幼体的氧化应激和细胞凋亡水平(n ≥ 3)。
(A)ROS 信号及量化结果。
(B)MitoSOX 信号及量化结果。
(C)MitoTracker 信号及量化结果。
(D)AO 染色的代表性图像和量化结果。EOM:浓度为 5 mg/L 的 EOM;BT:浓度为 5 μM 的甜菜碱。不同字母表示差异显著。
(3)EOM 通过 AHR 介导的 mettl14/ mettl3 下调抑制 m6A RNA 甲基化
如图 3A 所示,通过药物抑制剂 CH 或基因敲除抑制 AHR 的活性,可以抵消 EOM 降低的全局 m6A RNA 甲基化水平,这表明 AHR 介导了 EOM 诱导的 m6A RNA 甲基化变化。CH 已被用作斑马鱼胚胎和人类上皮细胞中的 AHR 拮抗剂。我们进一步检测了 m6A 调控因子(包括 METTL14、KIAA1429和 FTO)的 mRNA 表达水平。图 3B 显示,EOM 明显降低了 mettl14 和 Mettl3 的 mRNA 表达水平,补充 CH 后这些变化得以恢复。免疫荧光染色显示,CH或AHR敲除可拮抗EOM下调的Mettl14蛋白表达(图3C)。AHR 的敲除效率见。我们还发现,阴性对照 MO 对上述结果没有检测到影响。为了研究 AHR 是否直接抑制 mettl3 和 mettl14 的转录,我们检测了 mettl3 和 mettl14 的启动子序列。如图 3D所示,在这两个基因的启动子区域发现了多个可预测的 XRE 位点。此外,体内双荧光素酶报告实验表明,EOM能显著提高mettl3和mettl14野生型启动子的荧光素酶活性,而突变体样本的荧光素酶活性则没有提高(图3E)。

图 3. 72 hpf 时斑马鱼幼体 mRNA 和蛋白表达水平(n ≥ 3)
(A)m6A 抗体染色的代表性图像及量化结果。
(B)mRNA 表达水平。
(C)Mettl14 抗体染色的代表性图像及定量结果。
(D)mettl14 / mettl3 启动子区域的 XRE 位点和体内双荧光素酶报告结果。* ,突变位点;EOM:浓度为 5 mg/L 的 EOM;CH:浓度为 0.5 μM 的 CH223191;AHRMO:ahr2 ma啉寡核苷酸。不同字母表示差异具有统计学意义。
(E)双荧光素酶报告分析
(4)过表达 mettl14 或 mettl3 可减轻 EOM 诱导的 ROS 过度产生和细胞凋亡
如图 4A 和 B 所示,过表达 mettl14 可显著抵消 EOM 诱导的斑马鱼幼体心脏细胞内 ROS 和 mtROS 的产生。AO 染色显示,过表达 mettl14 可减轻 EOM 诱导的斑马鱼胚胎心脏凋亡(图 4C)。对裂解的caspase-3的免疫染色进一步证实了过表达mettl14对EOM诱导的心脏凋亡的拮抗作用(图4D)。同样,我们发现过表达 mettl3 也能抵消 EOM 诱导的细胞内 ROS/ mtROS 生成和斑马鱼幼体心脏凋亡(图 4E-G)。

图 4.过表达 mettl14/3 后 72 hpf 斑马鱼幼体的氧化应激和细胞凋亡水平(n ≥ 3)。
(A,E)ROS 信号及量化结果。
(B,F) MitoSOX 信号及量化结果。
(C,G)斑马鱼心脏 AO 染色的代表性图像及量化结果。
(D)对裂解的caspase-3的免疫染色
(E)Caspase-3裂解染色的代表性图像和定量结果。EOM:5 mg/L 的 EOM;Mettl14:mettl14 mRNA;Mettl3:mettl3 mRNA。不同字母表示差异显著。
(5) EOM 诱导全基因组 m6A RNA 甲基化变化
MeRIP-seq 结果显示,m6A 峰主要集中在中央编码区,并显著富集在终止密码子附近(图 5A)。EOM 在斑马鱼幼体心脏中诱导了广泛的全基因组 m6A RNA 甲基化变化,6815 个位点上调,918 个位点下调(图 5B)。富集分析显示,受影响的基因参与了包括神经嵴细胞迁移和心脏发育在内的重要生物学过程,ECMreceptor interaction、TGF-β、mTOR、Wnt 和 MAPK 信号通路是富集程度最高的通路之一(图 5C)。与仅添加EOM组相比,添加CH组进一步诱导了14,608个高甲基化位点和11,455个低甲基化位点(图6B)。我们重点研究了EOM诱导的m6A RNA甲基化异常变化,CH缓解了这些变化,我们认为这些变化的基因(如图5D所示,2509个基因位于Q1和Q4)是EOM诱导的心脏缺陷的罪魁祸首。我们注意到,除了 TGF-β 和 MAPK 信号通路外,细胞凋亡也在此时富集(图 5E,F)。图 6F 显示了两个与细胞凋亡相关的基因 traf4a 和 bbc3 的 m6A RNA 甲基化模式,我们选择了这两个基因作进一步研究。
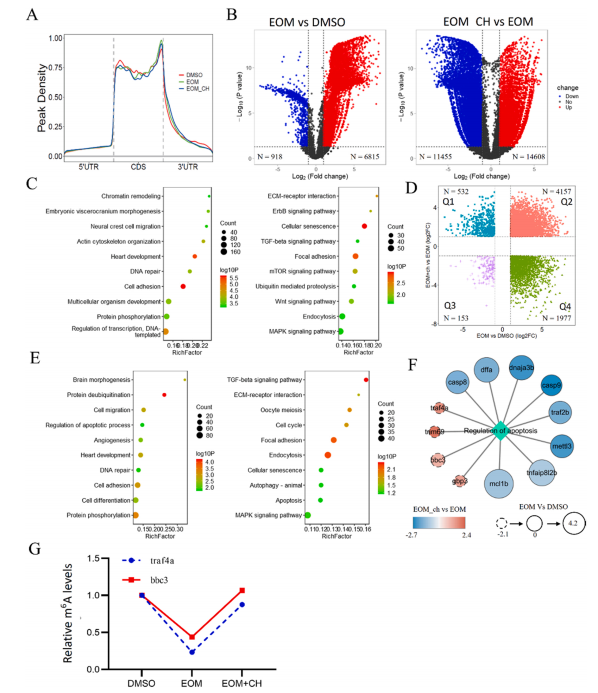
图 5.暴露于含或不含 CH 的 EOM 的斑马鱼幼体心脏中全基因组 m6A RNA 甲基化变化(n = 3)
(A)减少的 m6A 峰的分布。
(B)火山图显示 EOM 与 DMSO 和 EOM_CH 与 EOM 的不同甲基化位点。
(C)EOM 与 DMSO 中不同甲基化位点基因的富集分析。
(D)象限图反映了 EOM 和 EOM_CH 中差异甲基化基因的分析结果。
(E)Q1 和 Q4 中不同甲基化位点基因的富集分析。
(F)Q2 和 Q4 中存在差异甲基化位点的凋亡相关基因的 Newwork 图。
(G)重排 m6A RNA 甲基化水平。
(6) mettl14介导了EOM诱导的traf4a和bbc3的过表达
如图6A所示,EOM样本中traf4a和bbc3的mRNA表达水平均有所增加,但traf4a的表达量折减相当有限。通过强制表达 mettl14,EOM 上调的这两个基因的转录水平被降至对照水平。无独有偶,免疫荧光结果也显示,EOM上调了bbc3的蛋白表达水平,但随后又被mettl14的过表达所抵消(图6B)。

图 6.72 hpf 时斑马鱼幼体的相关基因表达水平(n ≥ 3)。
(A)traf4a 的 mRNA 表达水平。
(B)bbc3染色的代表性图像及量化结果。EOM:浓度为 5 mg/L 的 EOM。
(7)EOM诱导的氧化应激和细胞凋亡都需要traf4a和bbc3
我们首先发现,敲除traf4a或bbc3都会减轻EOM引起的贲门畸形。如图 7A 和 B 所示,在斑马鱼胚胎心脏中,敲除 traf4a 或 bbc3 均可终止 EOM 诱导的细胞内 ROS 和 mtROS 过度生成。此外,裂解天冬酶3免疫染色结果表明,敲除traf4a或bbc3可拮抗EOM诱导的斑马鱼胚胎心脏凋亡(图7C)。
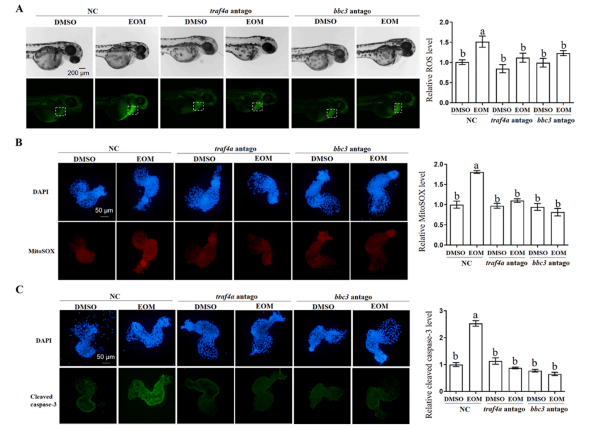
图 7.traf4a 或 bbc3 基因敲除后 72 hpf 斑马鱼幼体的氧化应激和细胞凋亡水平(n ≥ 3)。
(A)MitoSOX 信号和量化结果。
(B)被裂解的 caspase-3 染色的代表性图像和量化结果。EOM:5 mg/L的EOM;NC:非特异性对照抗原;不同字母表示差异有统计学意义
全文总结
结果表明,PM2.5的EOM通过AHR介导的mettl14下调诱导了广泛的m6A RNA甲基化变化,从而增加了traf4a和BBC3的表达水平,导致氧化应激和细胞凋亡,最终导致心脏发育异常(图8)。由于AHR拮抗剂和甲基供体如黄烷酮、姜黄素和甜菜碱广泛存在于日常食物中,它们可能是治疗PM2.5诱导的异常AHR激活和m6A RNA甲基化变化的良好候选者。
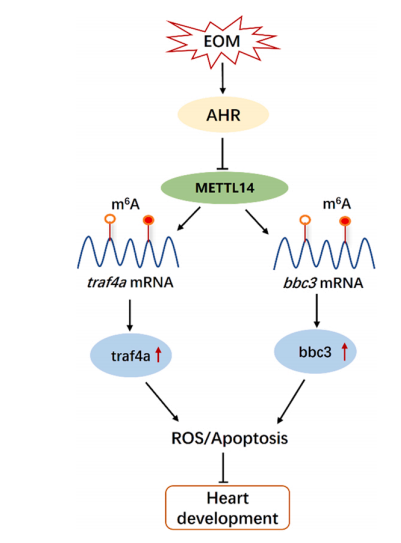
图8. 模型显示AHR介导的m6A RNA甲基化是PM2.5对心脏发育毒性的原因之一。
EOM 激活的 AHR 通过下调 mettl14 抑制 m6A RNA,进而提高 traf4a 和 bbc3 的表达水平,导致氧化应激和细胞凋亡,最终导致斑马鱼幼体心脏畸形。
参考文献
Ji C, Tao Y, Li X, Wang J, Chen J, Aniagu S, Jiang Y, Chen T. AHR-mediated m6A RNA methylation contributes to PM2.5-induced cardiac malformations in zebrafish larvae. J Hazard Mater. 2023 Sep 5;457:131749. doi: 10.1016/j.jhazmat.2023.131749. Epub 2023 Jun
云序生物m6A修饰研究五大模块
01 m6A RNA修饰测序
m6A RNA修饰测序(m6A-MeRIP-seq)
对m6A RNA甲基化,目前流行的检测手段为m6A-MeRIP-Seq技术,适用于m6A RNA甲基化谱研究,快速筛选m6A RNA甲基化靶基因。云序可提供mRNA和多种非编码RNA的m6A测序:
m6A 全转录组测序(涵盖mRNA,LncRNA,circRNA)
m6A LncRNA测序(涵盖LncRNA和mRNA)
m6A Pri-miRNA测序(涵盖Pri-miRNA和mRNA)
m6A mRNA测序
m6A miRNA测序
02检测整体m6A修饰水平
比色法检测整体RNA修饰水平
快速检测m6A整体甲基化水平
03 m6A RNA修饰上游酶的筛选
m6A RNA修饰相关酶PCR芯片
寻找上游直接调控m6A RNA甲基化的甲基转移酶。
04 m6A RNA修饰靶基因验证
MeRIP-qPCR/GenSeq® MeRIP试剂盒
云序提供各类不同修饰的MeRIP-qPCR服务以及销售GenSeq® MeRIP试剂盒,可针对mRNA,lncRNA,环状RNA等不同类型的RNA分子进行检测,低通量验证RNA修饰靶基因表达水平。
05机制互作研究
5.1 RIP-seq/qPCR/GenSeq® RIP试剂盒
筛选或验证RNA修饰直接靶点,研究RNA修饰靶基因的调控机制。
5.2 RNA pull down -MS/WB
筛选或验证目标RNA互作基因或蛋白,研究相应的分子调控机制。
5.3 ChIP-seq
筛选或验证目标蛋白与DNA互作,研究相应的分子调控机制。
5.4 Ribo-seq
筛选翻译水平发生变化的RNA分子,研究相应的分子调控机制。
5.5 SLAM-seq
筛选稳定性发生变化的RNA分子,研究相应的分子调控机制。
5.6 CHIRP-seq
筛选或验证目标RNA与DNA互作,研究相应的分子机制。
云序生物服务优势
优势一:云序累计支持客户发表100 +篇RNA修饰SCI论文,合计影响因子1000 +
优势二:累计完成数千例 RNA甲基化测序样本,全面覆盖医口、农口等各类样本。
优势三:全面检测mRNA和各类非编码RNA(circRNA,lncRNA,Pri-miRNA等)。
优势四:提供m6A一站式服务:m6A整体水平检测、m6A-MeRIP-seq、MeRIP-qPCR验证、RIP和RNA pull-down、Ribo-seq、SLAM-seq、CHIRP-seq、CHIP-seq等。
优势五:率先研发超微量MeRIP测序技术,RNA量低至500ng起。
优势六:国内全的RNA修饰测序平台,提供m6A、m5C、m1A、m7G、m3C、O8G、ac4C乙酰化和2'-O-甲基化、假尿嘧啶RNA测序。
云序生物m6A多组学一站式服务

云序客户RNA修饰部分文章列表

相关产品
m6A RNA甲基化测序
m5C RNA甲基化测序
m1A RNA甲基化测序
m7G RNA甲基化测序
ac4C RNA乙酰化测序
O8G RNA氧化修饰测序
2’-O-RNA甲基化测序
m6Am RNA甲基化测序
假尿嘧啶测序
RNA pulldown
RNA-seq
RIP测序
Ribo-seq
SLAM-seq
CHIRP-seq
CHIP-seq
往期回顾
云序客户m6A高分文章|揭示组蛋白乙酰化与m6A修饰在眼部黑色素瘤发生中的共同作用机制
用户文章 IF=19.2|范先群院士团队利用m1A MeRIP-seq联合多组学测序揭示眼黑色素瘤调控新机制
云序客户| m6A MeRIP-seq助力揭示早发糖尿病表观调控新机制
客户文章|IF=10.6—甲状腺癌中m5C甲基转移酶NSUN2通过维持tRNA的稳定性促进密码子依赖性的致癌翻译
北大伊成器团队Nature Reviews重磅发文:非m6A热门修饰调控与功能一文速览!
云序客户| MeRIP-seq+RIP-seq技术揭示 m6A 甲基化调控BMSCs成骨分化新机制
客户文章| Nature子刊 揭示了FMR1通过m6A修饰调控早期胚胎发育的分子机制
用户文章m6A专题|IF=9.8|m6A去甲基化酶ALKBH5缺乏会加重钴致神经退行性损伤
1区,IF=27| 云序m6A MeRIP-seq助力鳞状细胞癌机制研究!
用户文章 1区 lncRNA m6A甲基化测序助力人脂肪干细胞成骨分化的调控机制研究
客户文章|1区,IF=9.995|m6A甲基化测序助力宫颈癌相关HPV病毒研究
用户文章IF=19.568|m5C修饰测序助力NSUN2调控病毒I型干扰素反应的机制研究
Nature 新发现:线粒体 m5C 修饰竟是肿瘤转移的元凶!
用户文章IF=14.9 1区:ac4C乙酰化调节人胚胎干细胞的自我更新
云序用户农口IF20+论文:植物mRNA存在ac4C新型修饰,对RNA稳定性及翻译产生重要影响
用户文章:m7G 甲基化参与调节心肌细胞增殖|新型RNA修饰研究
客户文章|新型RNA修饰之m7G揭示急性髓系白血病发病机制
云序用户植物文章6连发:植物当中各类RNA甲基化测序该怎么发?
庆祝云序用户m1A、m6A、m5C RNA甲基化测序文章三连发