

产品分类
公司荣誉

公司图片


联系方式
公司新闻
《Cell》新发现!m6A-MeRIP-seq技术揭示m6A 修饰新功能!
发布时间:2021-08-02 13:50 | 点击次数:
2021年4月29日日内瓦大学Ramesh S. Pillai、David Homolka研究组合在著名学术期刊《Cell》在线发表了一项新成果,该团队发现剪接位点m6A甲基化通过阻止剪接因子U2AF35与前体mRNA结合进而抑制RNA剪接,结合后续的功能机制研究,揭示了m6A甲基化修饰新的调控机制。接下来,就让我们一起来看下研究人员是如何利用高通量测序技术RNA-seq和m6A-MeRIP-seq揭示m6A甲基化修饰调控RNA剪接这一新功能的。

发表日期:2021年4月29日
影响因子:41.582
研究方法::m6A meRIP-seq、RNA-seq、LC-MS/MS等
文章链接:https://pubmed.ncbi.nlm.nih.gov/33930289/
文章概要
6-methyladenosine (m6A) RNA修饰被广泛用于改变mRNA的命运。本文中作者证明了秀丽隐杆线虫的甲基化修饰蛋白METT-10(小鼠METTL16的同源蛋白)沉积在S-腺苷甲硫氨酸(SAM)合成酶前体mRNA的3’剪接位点(AG)上的一个m6A修饰,可抑制其正确剪接和蛋白产生。这种机制是由丰富的饮食触发的,以m6A介导的开关来停止SAM的产生并调节其内稳态。虽然哺乳动物SAM合成酶pre-mRNA不受此机制调控,但3’剪接位点m6A修饰抑制剪切在哺乳动物中是保守的。该修饰通过物理方式阻止必要的剪接因子U2AF35识别3’剪接位点。作者认为使用剪切位点m6A修饰是一种古老的剪接调节机制。
研究思路

研究内容
一、秀丽隐杆线虫的m6A转录组
已知哺乳动物中主要有两类m6A“编写器”:METTL3/METTL14复合物以及METTL16,为了研究 METTL16 催化活性的保守作用,作者选择了秀丽隐杆线虫(以下称为蠕虫)。蠕虫直系同源物 METT-10包含高度保守的 RNA 甲基转移酶域,但缺乏哺乳动物 METTL16 中发现的 VCR域(图 1A)。通过LC-MS/MS技术检测来自成虫的总 RNA 和 poly(A)+ RNA 中的各种核糖和碱基修饰发现,在来自所有三种生物来源的 poly(A)+ RNA 中检测到 m6 A 修饰,包括秀丽隐杆线虫(图 1B)。为了鉴定携带 m6A 甲基化的蠕虫转录本,作者使用来自成年秀丽隐杆线虫的 poly(A)+ RNA 和小鼠睾丸 RNA (阳性对照)的混合物进行了 m6A-meRIP-seq(图 1C)与超过 20,000 个小鼠峰相比,在蠕虫 poly(A)+ 转录组中仅鉴定出 176 个 m6A 峰(图 1D-E),这可能是由于蠕虫中不存在 METTL3/METTL14 “编写器”复合体。

在确认蠕虫 poly(A)+ RNA上存在 m6A 后,作者接下来寻找METT-10甲基化靶标,首先使用了m6A-meRIP-seq数据集的比较分析来鉴定 poly(A)+ 转录本,这些转录本在METT-10 敲除 (KO) 突变体中显示m6A 甲基化减少(图 2F)。其中,前 20 个编码 U6snRNA 序列(图2G)。与此一致,对总 RNA 样本进行的单独 m6A-meRIP-seq实验显示 U6 snRNA 读数的富集程度更高(图 2H)。已有研究表明人类 U6 snRNA 在九聚体基序 (UACm6AGAGAA) 内被人 METTL16 甲基化,在此m6A-meRIP-seq读数的映射显示蠕虫 U6 snRNA 也在相同的位点(图2I)。重要的是,这个 m6A 信号在 METT-10 KO 突变体中消失(图 2H 和2I)。作为独立验证,通过 SCARLET 方法(以核苷酸特异性方式检测甲基化状态),对成虫总 RNA 的分析证实了甲基化共有基序中这种特定腺苷(A)的甲基化(图2J)。此外,在 METT-10 KO 中显示 m6A水平显着下降的其他转录本是 sams-3、sams-4 和 sams-5(图2G),这些重复的基因编码 SAM 合成酶,该酶负责产生甲基供体 SAM,这是细胞中甲基化反应所必需的。综上,作者将蠕虫 METT-10 鉴定为 m6A RNA 甲基转移酶,并且与其哺乳动物直系同源物 METTL16 一样,它具有 U6 snRNA 和 SAM 合成酶 RNA两个保守靶点。

作者通过将m6A-meRIP-seq数据集映射到蠕虫基因组,在sams m6A峰上发现了一个甲基化基序(UACm6AGAAAC),其中该基序中的甲基化腺苷位于内含子2的3’剪接位点(AG)(图3A)。除了sams-3/4转录本的成熟剪接蛋白编码型(PC亚型),作者还检测到了两个未使用内含子2内3’剪接位点的非编码型:使用上游隐藏的3’剪接位点的替代剪接变异体(AS亚型) 和内含子保留型(IR亚型) (图3C)。其中,与野生型(WT)蠕虫相比,METT-10 KO中内含子2的总体含量较低,表明在没有3‘剪接位点甲基化时,其有效剪接(图3B)。与此一致的是,RNA-seq数据显示,METT-10 KO中优先使用3’剪接位点(产生PC亚型) (图3C)。这种m6A介导的剪接抑制的结果是METT-10 KO中sams mRNA水平普遍增加(图3D)。综上所述,蠕虫通过METT10介导的3’剪接位点m6A甲基化来抑制SAM合成酶mRNA的剪接和产生。
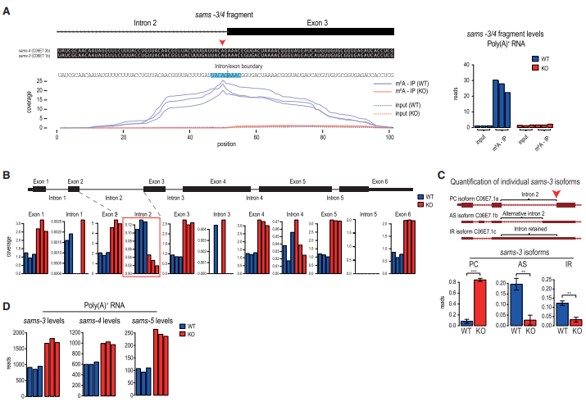
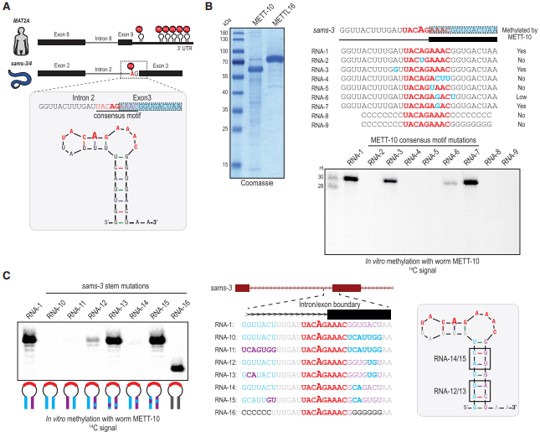
二级结构预测显示,跨越3’剪接位点的sams-3 pre-mRNA的30-nt RNA片段折叠成茎环结构,甲基化共识基序(UACm6AGAAAC)占据了部分环区(图4A)。为了证实该序列可以被蠕虫METT-10甲基化,作者将这30-nt RNA与重组全长蠕虫METT-10和放射性14C-SAM作为甲基供体孵育,发现RNA在3’剪接位点(AG)特异性甲基化,其中腺苷、共识基序内的单一或三重突变消除了METT-10的体外甲基化活性(图4B)。茎秆区域对甲基化也很关键,将茎环区域碱基突变后失去甲基化活性 (图4C)。这些结果表明,sams pre-mRNA的甲基化共识基序和3’剪接位点的茎环形成是其被METT-10识别的先决条件。
五、蠕虫通过甲基化SAM合成酶转录子的3’剪接位点下调SAM合成酶的表达以响应高营养饮食
上述实验3’剪接位点甲基化介导的剪接抑制,蠕虫是在高营养琼脂培养基上生长的,而将饮食改为低营养琼脂平板后导致WT和METT-10 KO蠕虫失去了这种剪接调控和类似的亚型表达模式(图5A)。为了直接确定剪接位点m6A甲基化对饮食变化的响应,作者用来自WT和METT-10 KO蠕虫的poly(A)+ RNA进行了m6A-meRIP-seq实验,这些poly(A)+ RNA来自两种不同的饮食,发现生长在高营养培养皿中的WT蠕虫sams-3 pre-mRNA内含子2的3’剪接位点发生强m6A甲基化修饰,而低营养培养皿中甲基化显著降低(图5B)。因此,3'剪接位点m6A甲基化是对高营养饮食的反应,以抑制SAM合成酶pre-mRNA适当的剪接和表达。由于RNA甲基化依赖于SAM作为甲基供体,作者还研究了该途径是否通过反馈抑制来调节细胞SAM水平。代谢组学分析显示,尽管WT蠕虫能够控制SAM水平,但METT10 KO蠕虫却不能,METT-10的丢失破坏了这种稳态,导致两种饮食条件下SAM浓度升高 (图5E)。通过WB实验检测了该酶的蛋白水平,与RNA分析一致,高营养饮食条件下的SAMS-3蛋白水平下降 (图5F)。因此,m6A修饰调控了蠕虫SAM内稳态。

六、 m6A甲基化阻止了必需剪接因子U2AF35对剪接位点的识别
在后生动物中,剪接因子对pre-mRNA内的关键顺式元件的识别是剪接起始的关键。U2AF是由U2AF35和U2AF65亚基组成的异二聚体,其中U2AF35已被证明直接结合AG二核苷酸的3‘剪接位点,因此作者想研究3’剪接位点甲基化是否会阻碍U2AF35的结合。等温量热法(ITC)实验显示,U2AF35与未甲基化的RNA相互作用强烈,但3’剪接位点m6A的存在使其亲和力降低了一个量级。因此,3’剪接位点m6A修饰通过物理上阻止必需剪接因子U2AF35对其的识别进而抑制pre-mRNA的正确剪接。
那么,m6A介导的抑制通路在哺乳动物系统中是否也存在呢?作者将基于蠕虫sams-3的转基因报告子构建体转染到人HeLa细胞中,通过RT-PCR实验发现与在蠕虫中表达相同构建体时有着相似的剪接模式,且甲基化共识基序突变体AS亚型水平降低,发生有效剪接(图7A)。为了证明剪接调控是由于m6A修饰的存在,在3’剪接位点人工引入了一个m6A,并进行体外剪接试验。发现未甲基化底物正常剪接,而3’剪接位点m6A修饰的底物显示缺少套索中间体和成熟剪接产物 (图7B)。已知METTL16缺失会导致小鼠植入前胚胎死亡,为了检测其催化活性的体内相关性,创建了一个携带催化基序突变体的基因敲入小鼠,发现纯合子催化活性和RNA结合活性丢失的METTL16突变体表现出发育致死率(图7C)。此外,METTL16条件性缺失(cKO)的小鼠会导致睾丸萎缩和生殖细胞发育受阻(图7D-E)。以上实验揭示了哺乳动物中3’剪接位点m6A甲基化的剪接抑制是保守的,以及METTL16催化活性在小鼠发育过程中的重要作用。

文章小结
文章通过m6A meRIP-seq、RNA-seq以及LC-MS/MS等技术揭示了3ʹ剪接位点的m6A 甲基化通过阻止必需剪接因子U2AF35对RNA的识别,从而抑制线虫和哺乳动物中的前体mRNA 剪接。在蠕虫中,这种机制用于调节剪接以响应饮食的变化。m6A修饰参与调控多种生物学功能,其调控机制异常还可能导致相关疾病或癌症的发生,本文3ʹ剪接位点m6A 甲基化剪接调控机制的发现将有助于以后对这些相关生物学功能或疾病的探索。
云序生物m6A修饰研究五大模块
01 m6ARNA修饰测序
m6A RNA修饰测序(m6A-meRIP-seq)
对m6A RNA甲基化,目前流行的检测手段为m6A-Seq技术,适用于m6A RNA甲基化谱研究,快速筛选m6A RNA甲基化靶基因。云序可提供mRNA和多种非编码RNA的m6A测序:
● m6A 全转录组测序(涵盖mRNA,LncRNA,circRNA)
● m6A LncRNA测序(涵盖LncRNA和mRNA)
● m6A Pri-miRNA测序(涵盖Pri-miRNA和mRNA)
● m6A mRNA测序
● m6A miRNA测序
02 检测整体m6ARNA修饰水平
LC-MS/MS检测整体RNA修饰水平
精准高效,可以实现一次检测,9类修饰水平检测,一步到位。
比色法检测整体RNA修饰水平
快速检测m6A整体甲基化水平
03 m6A RNA修饰上游酶的筛选
m6A RNA修饰相关酶PCR芯片
寻找上游直接调控m6A RNA甲基化的甲基转移酶。
04 m6A RNA修饰靶基因验证
meRIP-qPCR
云序提供各类不同修饰的meRIP-qPCR服务,可针对mRNA,lncRNA,环状RNA等不同类型的RNA分子进行检测,低通量验证RNA修饰靶基因表达水平。
05机制互作研究
5.1 RIP-seq/qPCR
筛选或验证RNA修饰直接靶点,研究RNA修饰靶基因的调控机制。
5.2 RNA pull down -MS/WB
筛选或验证目标RNA互作基因或蛋白,研究相应的分子调控机制。
5.3 双荧光素酶实验
验证两基因互作,研究相应的分子调控机制。
5.4 ChIP-seq
筛选或验证目标蛋白与DNA互作,研究相应的分子调控机制。
云序生物服务优势
优势一:发表10分以上文章最多的m6A RNA甲基化测序服务平台。云序已累计支持客户发表52+篇高水平文章,合计影响因子450分+,是国内支持发文最多、累计影响因子最高的公司。
优势二:至今完成4000+例m6A测序样本,全面覆盖医口、农口等各类样本。
优势三:全面检测mRNA和各类非编码RNA(circRNA,lncRNA,Pri-miRNA等)。
优势四:提供m6A一站式服务:m6A整体水平检测、m6A测序、MeRIP-qPCR验证、RIP和RNA pull-down等。
优势五:率先研发超微量MeRIP测序技术,RNA量低至500ng起。
优势六:国内最全的RNA修饰测序平台,提供m6A、m5C、m1A、m7G、m3C、O8G、ac4C乙酰化和2'-O-甲基化测序。
云序客户RNA修饰部分文章列表

相关产品
m1A RNA甲基化测序m5C RNA甲基化测序
m7G (m3C)RNA 甲基化测序
2’-O-RNA氧化测序
ac4C乙酰化测序
o8G RNA氧化测序
比色法/LC-MS检测整体m6A甲基化水平
RNA修饰相关酶PCR芯片
ATAC-seq
ChIP-seq
往期回顾
经典思路|云序客户circRNA+m6A案例教您快速发表5分文章
m6A项目文章|ALKBH5通过调控WNT5A稳定性促进缺血后血管生成
云序客户再发15分文章:FBW7靶向m6A结合蛋白YTHDF2抑制卵巢癌
Science新发现 |RNA以m6A依赖方式调节染色质状态和基因转录
NatCommun | 云序客户揭示Mettl3协调癌症的生长和转移的分子机制
云序客户余健秀课题组m6A方向再次取得重大发现——SUMO化促进YTHDF2结合m6A修饰的mRNA影响癌症进展
项目文章|Nature子刊m6A修饰携手miRNA揭示脱氧胆酸在胆囊癌中作用机制